Molecular Effects of Xylella fastidiosa and Drought Combined Stress in Olive Trees

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
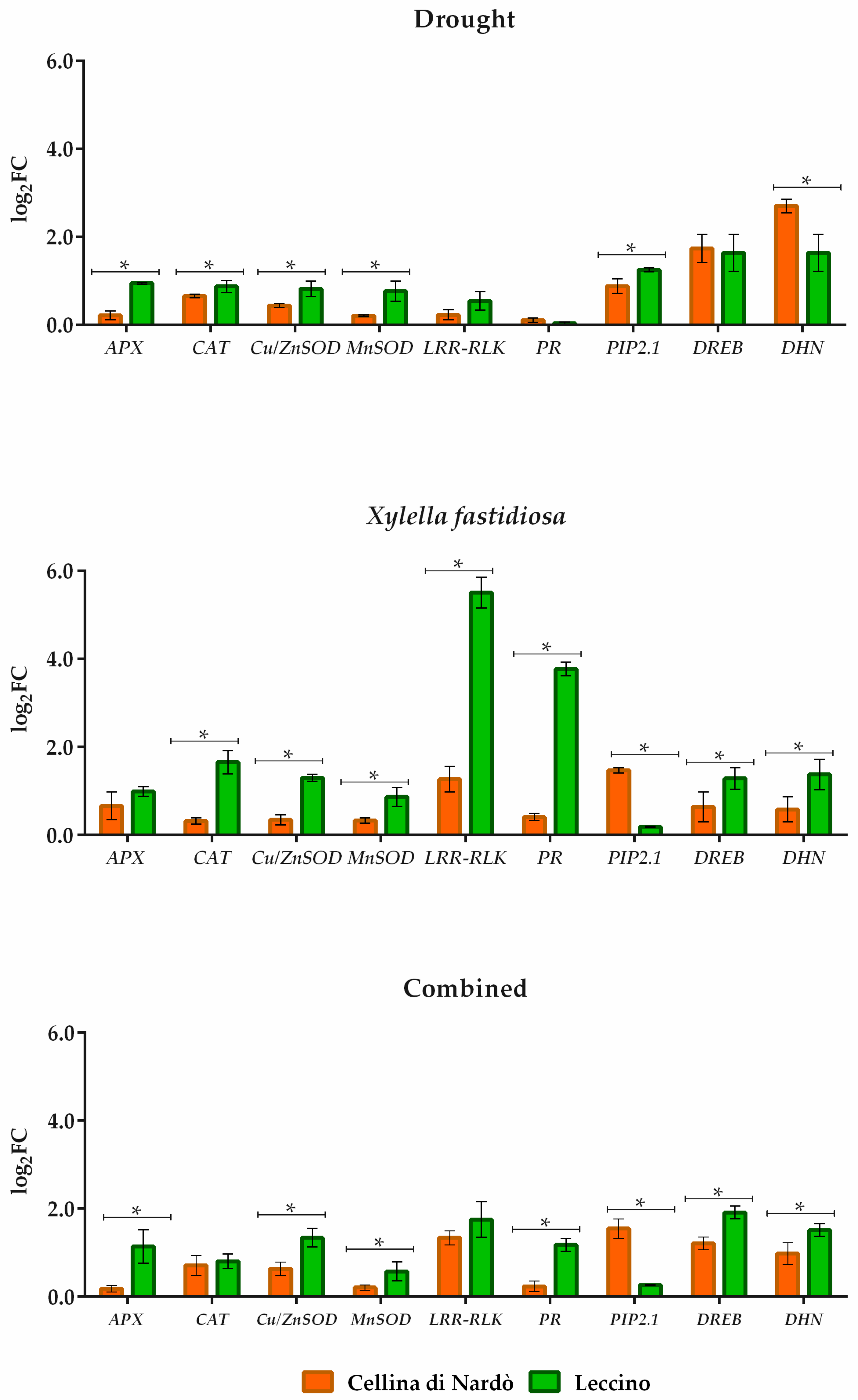
2. Results
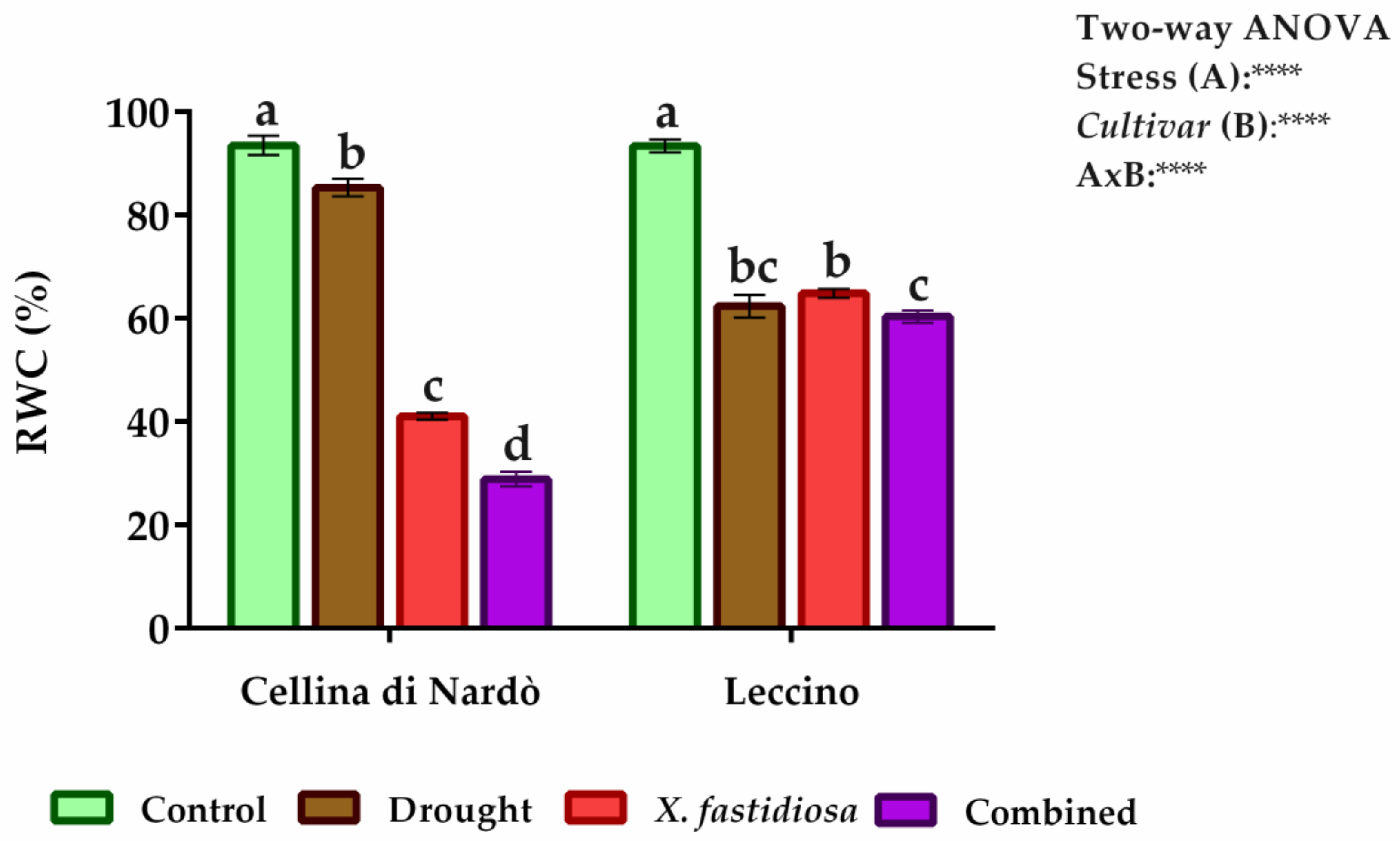
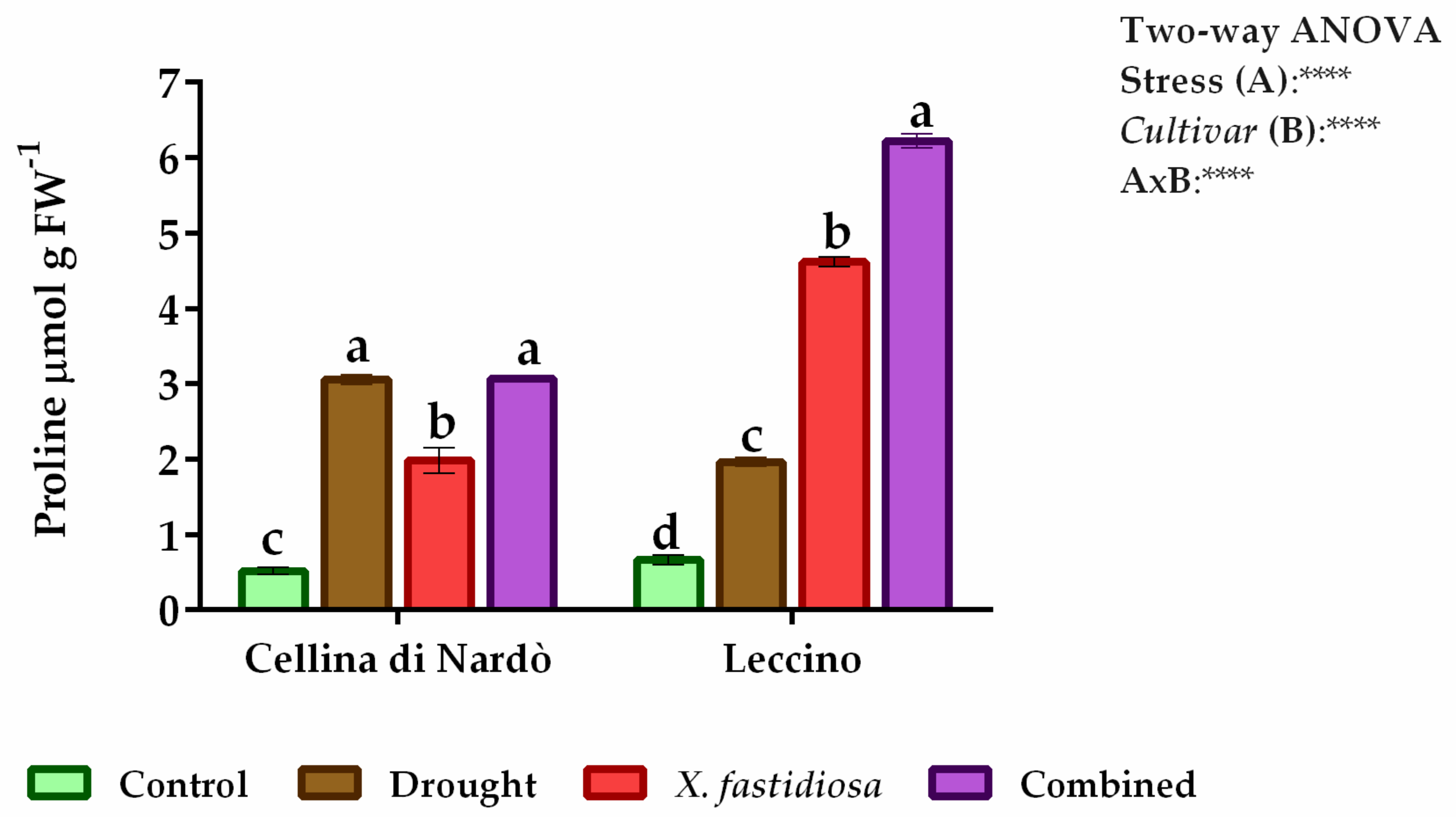
2.1. Estimation of Water and Proline Content and Enzyme Assays
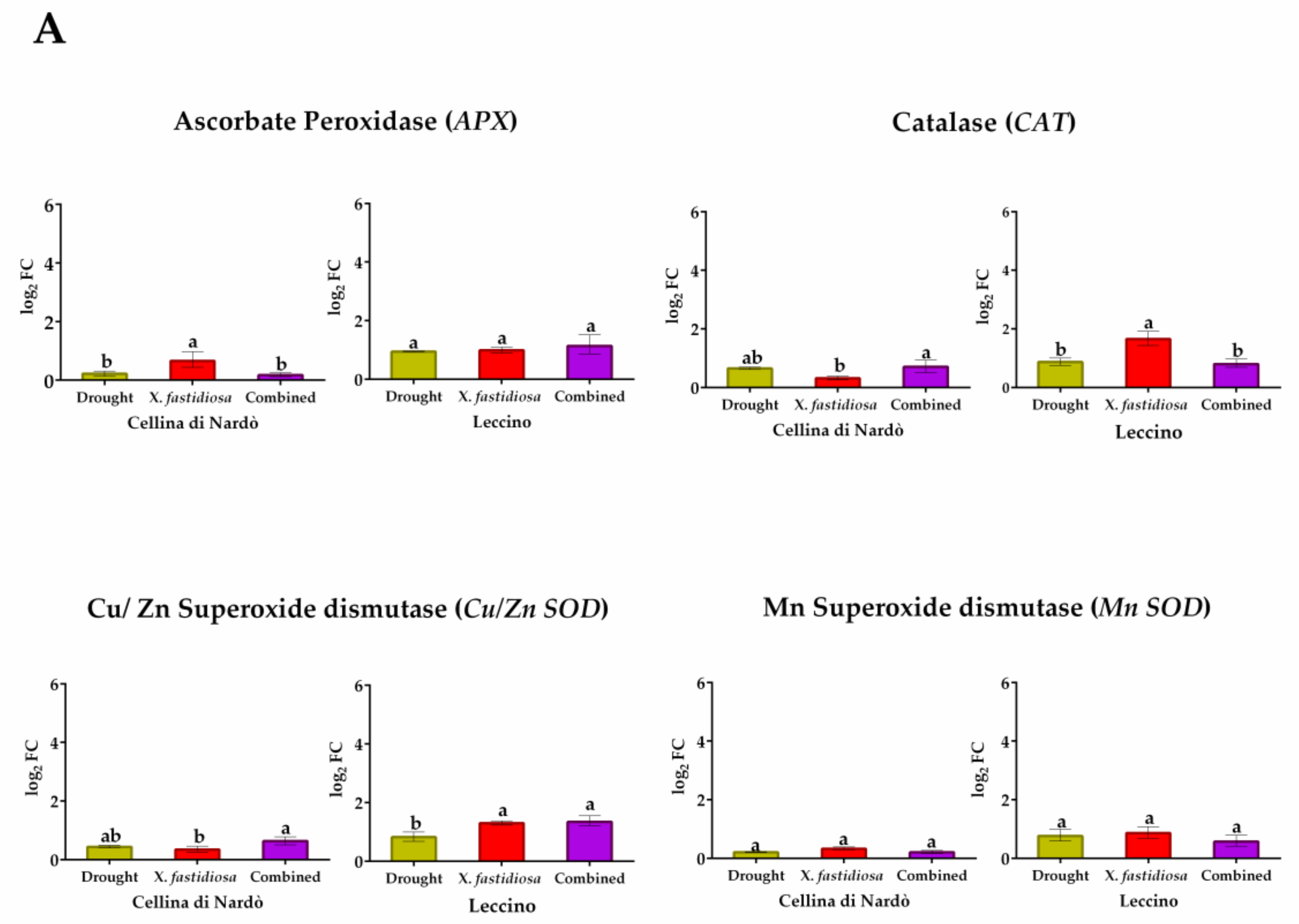
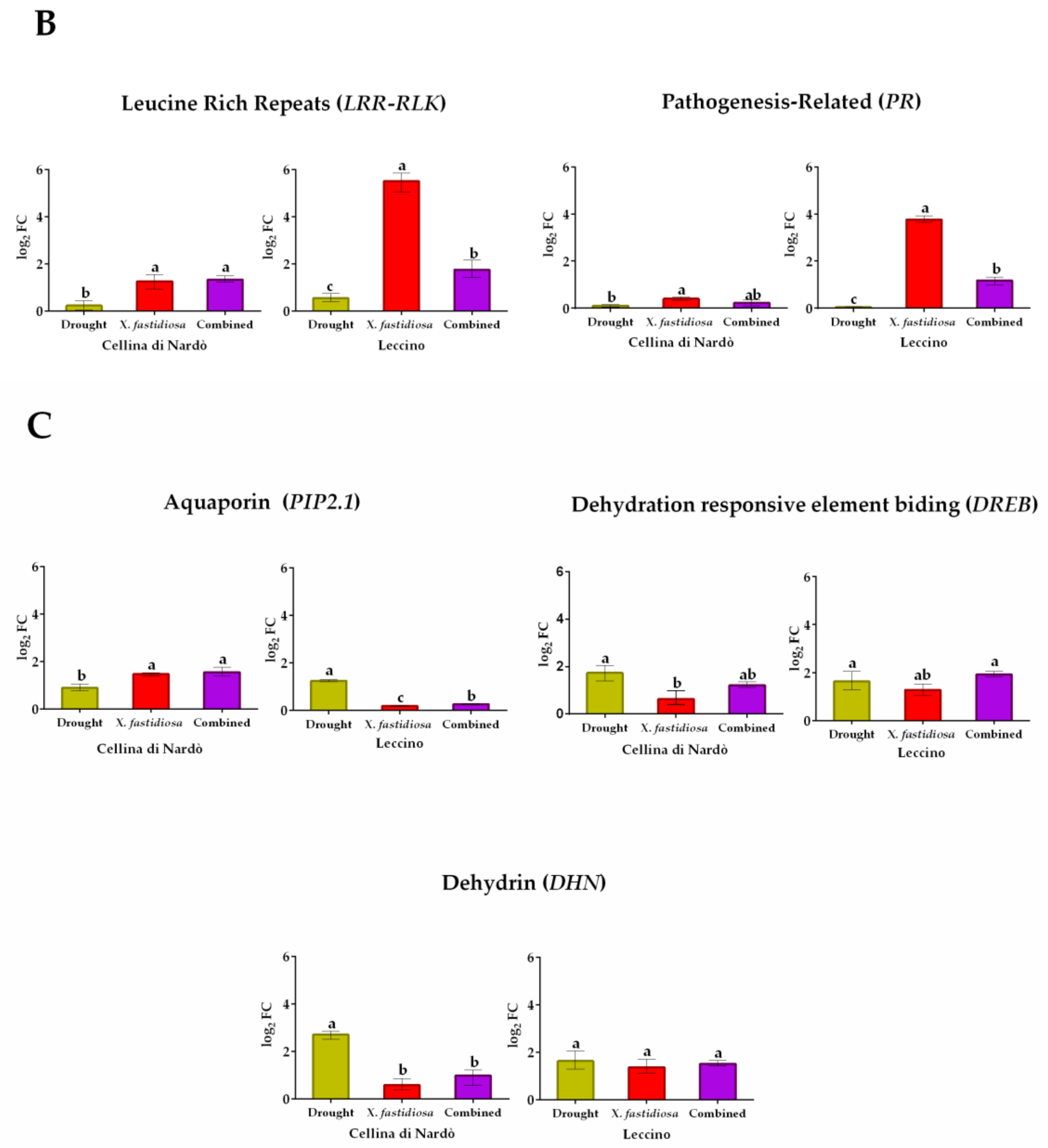
2.2. Gene Expression Analysis under Individual and Combined Stresses
3. Discussion
4. Materials and Methods
4.1. Field Conditions and Plant Material
4.2. Relative Water Content Measurement
4.3. Free Proline Determination
4.4. Antioxidant Enzymes Determinations
4.5. Total RNA Isolation, cDNA Synthesis, and Real-Time PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abuqamar, S.; Luo, H.; Laluk, K.; Mickelbart, M.V.; Mengiste, T. Crosstalk between biotic and abiotic stress responses in tomato is mediated by the AIM1 transcription factor. Plant J. 2009, 58, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Mengiste, T.; Chen, X.; Salmeron, J.; Dietrich, R. The Botrytis susceptible1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell 2003, 15, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Mittler, R. Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c. Plant Physiol. 2005, 139, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Narsai, R.; Wang, C.; Chen, J.; Wu, J.; Shou, H.; Whelan, J. Antagonistic, overlapping and distinct responses to biotic stress in rice (Oryza sativa) and interactions with abiotic stress. BMC Genomics. 2013, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Shaik, R.; Ramakrishna, W. Genes and co-expression modules common to drought and bacterial stress responses in Arabidopsis and rice. PLoS ONE 2013, 8, 77261. [Google Scholar] [CrossRef] [PubMed]
- Shaik, R.; Ramakrishna, W. Machine learning approaches distinguish multiple stress conditions using stress-responsive genes and identify candidate genes for broad resistance in rice. Plant Physiol. 2014, 164, 481–495. [Google Scholar] [CrossRef]
- Sharma, R.; Vleesschauwer, D.D.; Sharma, M.K.; Ronald, P.C. Recent advances in dissecting stress-regulatory crosstalk in rice. Mol. Plant 2013, 6, 250–260. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Lilley, C.J.; Urwin, P.E. Identification of genes involved in the response of Arabidopsis to simultaneous biotic and abiotic stresses. Plant Physiol. 2013, 162, 2028–2041. [Google Scholar] [CrossRef]
- Bostock, R.M.; Pye, M.F.; Roubtsova, T.V. Predisposition in plant disease: Exploiting the nexus in abiotic and biotic stress perception and response. Annu. Rev. Phytopathol. 2014, 52, 517–549. [Google Scholar] [CrossRef]
- Kissoudis, C.; Van de Wiel, C.; Visser, R.G.F.; Van Der Linden, G. Enhancing crop resilience to combined abiotic and biotic stress through the dissection of physiological and molecular crosstalk. Front. Plant Sci. 2014, 5, 207. [Google Scholar] [Green Version]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef] [PubMed]
- Prasch, C.M.; Sonnewald, U. Signaling events in plants: Stress factors in combination change the picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol. 2013, 162, 1849–1866. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Barah, P.; Suarez-Rodriguez, M.C.; Bressendorf, S.; Friis, P.; Costantino, P. Transcriptome responses to combinations of stresses in Arabidopsis. Plant Physiol. 2013, 161, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Mattson, W.J.; Haack, R.A. The role of drought stress in provoking outbreaks of phytophagous insects. In Insect Outbreaks; Barbosa, P., Schultz, J.C., Eds.; Academic Press: New York, NY, USA, 1987; pp. 365–407. [Google Scholar]
- Choi, H.K.; Iandolino, A.; Silva, F.G.; Cook, D.R. Water deficit modulates the response of Vitis vinifera to the Pierce’s disease pathogen Xylella fastidiosa. Mol. Plant-Microbe Interact. 2013, 26, 643–657. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M.; Ishiga, Y.; Kaundal, A.; Udayakumar, M.; Mysore, K.S. Drought stress acclimation imparts tolerance to Sclerotinia sclerotirum and Pseudomonas syringae in Nicotiana benthamiana. Int. J. Mol. Sci. 2013, 14, 9497–9513. [Google Scholar] [CrossRef]
- Gupta, A.; Sarkar, A.K.; Senthil-Kumar, M. Global transcriptional analysis reveals unique and shared response in Arabidopsis thaliana exposed to combined drought and pathogen stress. Front. Plant Sci. 2016, 7, 686. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. The reactive oxygen gene network in plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Chiappetta, A.; Muto, A.; Bruno, L.; Woloszynska, M.; Vanlijsebettens, M.; Bitonti, M.B. A dehydrin gene isolated from feral olive enhances drought tolerance in Arabidopsis transgenic plants. Front. Plant Sci. 2015, 6, 392. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (southern Italy). J. Plant Pathol. 2013, 95, 668. [Google Scholar]
- Luvisi, A.; Aprile, A.; Sabella, E.; Vergine, M.; Nicolì, F.; Nutricati, E.; Miceli, A.; Negro, C.; De Bellis, L. Xylella fastidiosa subsp. pauca (CoDiRO strain) infection in four olive (Olea europaea L.) cultivars: Profile of phenolic compounds in leaves and progression of leaf scorch symptoms. Phytopathol. Mediterr. 2017, 56, 259–273. [Google Scholar]
- Sabella, E.; Luvisi, A.; Aprile, A.; Negro, C.; Vergine, M.; Nicolì, F.; Miceli, A.; De Bellis, L. Xylella fastidiosa induces differential expression of lignification related-genes and lignin accumulation in tolerant olive trees cv. Leccino. J. Plant Physiol. 2018, 220, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, G.; Saponari, M.; Boscia, D.; D’Attoma, G.; Morelli, M.; Martelli, G.; Almeida, R.P.P. Intercepted isolates of Xylella fastidiosa in Europe reveal novel genetic diversity. Eur. J. Plant Pathol. 2016, 146, 85–94. [Google Scholar] [CrossRef]
- Cardinale, M.; Luvisi, A.; Meyer, J.B.; Sabella, E.; De Bellis, L.; Cruz, A.C.; Ampatzidis, Y.; Cherubini, P. Specific Fluorescence in Situ Hybridization (FISH) Test to Highlight Colonization of Xylem Vessels by Xylella fastidiosa in Naturally Infected Olive Trees (Olea europaea L.). Front. Plant Sci. 2018, 9, 431. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef]
- Bosso, L.; Febbraro, M.; Cristinzio, G.; Zoina, A.; Russo, D. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Keller, R.P.; Lodge, D.M.; Finnoff, D.C. Risk assessment for invasive species produces net bioeconomic benefits. Proc. Natl. Acad. Sci. USA 2007, 104, 203–207. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Principles Governing IPCC Work. Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2013; Volume 2, Available online: https://www.ipcc.ch/site/assets/uploads/sites/2/2019/05/SR15_Chapter1_Low_Res.pdf (accessed on 21 October 2019).
- Ramegowda, V.; Senthil-Kumar, M.; Udayakumar, M.; Kirankumar, S.M. A high-throughput virus induced gene silencing protocol identifies genes involved in multistress tolerance. BMC Plant Biol. 2013, 13, 193. [Google Scholar]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Zarattini, M.; Forlani, G. Toward unveiling the mechanisms for transcriptional regulation of proline biosynthesis in the plant cell response to biotic and abiotic stress conditions. Front. Plant Sci. 2017, 8, 927. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Riedel, H.; Saw, N.M.M.T.; Kütük, O.; Mewis, I.; Jäger, H.; Knorr, D.; Smetanska, I. Effects of pulsed electric field on secondary metabolism of Vitis vinifera L. cv. Gamay Fréaux suspension culture and exudates. Appl. Biochem. Biotechnol. 2011, 164, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Chen, M.; Su, Y.; Wu, N.; Yuan, M.; Yuan, S.; Brestic, M.; Zivcak, M.; Zhang, H.; Chen, Y. Comparison on photosynthesis and antioxidant defense systems in wheat with different ploidy levels and octoploid triticale. Int. J. Mol. Sci. 2018, 19, 3006. [Google Scholar] [CrossRef] [PubMed]
- Nicolì, F.; Negro, C.; Nutricati, E.; Vergine, M.; Aprile, A.; Sabella, E.; Damiano, G.; De Bellis, L.; Luvisi, A. Accumulation of azelaic acid in Xylella fastidiosa-infected olive trees: A mobile metabolite for health screening. Phytopathology 2018, 109, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, P.H.; DeVay, J.E.; Meredith, C.P. Roles of water stress and phytotoxins in the development of Pierce’s disease of the grapevine. Physiol. Mol. Plant Pathol. 1988, 32, 1–15. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant responses to water stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Lugojan, C.; Ciulca, S. Evaluation of relative water content in winter wheat. J. Hortic. For. Biotechnol. 2011, 15, 173–177. [Google Scholar]
- Solla, A.; Gil, L. Influence of water stress on Dutch elm disease symptoms in Ulmus minor. Can. J. Bot. 2002, 80, 810–817. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Genga, A.; Siciliano, T.; Nutricati, E.; Nicolì, F.; Vergine, M.; Negro, C.; De Bellis, L.; Luvisi, A. Xylem cavitation susceptibility and refilling mechanisms in olive trees infected by Xylella fastidiosa. Sci. Rep. 2019, 9, 9602. [Google Scholar] [CrossRef]
- Hu, L.; Wang, Z.; Du, H.; Huang, B. Differential accumulation of dehydrins in response to water stress for hybrid and common bermudagrass genotypes differing in drought tolerance. J. Plant Physiol. 2010, 167, 103–110. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Secchi, F.; Lovisolo, C.; Schubert, A. Expression of OePIP2.1 aquaporin gene and water relations of Olea europaea twigs during drought stress and recovery. Ann. Appl. Biol. 2007, 150, 163–167. [Google Scholar] [CrossRef]
- Rapicavoli, J.N.; Blanco-Ulate, B.; Muszyński, A.; Figueroa-Balderas, R.; Morales-Cruz, A.; Azadi, P.; Dobruchowska, J.M.; Castro, C.; Cantu, D.; Roper, M.C. Lipopolysaccharide O-antigen delays plant innate immune recognition of Xylella fastidiosa. Nat. Commun. 2018, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Roper, C.; Castro, C.; Ingel, B. Xylella fastidiosa: Bacterial parasitism with hallmarks of commensalism. Curr. Opin. Plant Biol. 2019, 50, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Yadav, S.; Wani, A.S.; Irfan, M.; Alyemini, M.N.; Ahmad, A. Impact of sodium nitroprusside on nitrate reductase, proline and antioxidant system in Solanum lycopersicum under salinity stress. Hort. Environ. Biotechnol. 2012, 53, 362–367. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Ben Rouinab, B.; Sensoyc, S.; Boukhrisa, M.; Ben Abdallah, F. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Xu, F.J.; Jin, C.W.; Liu, W.J.; Zhang, Y.S.; Lin, X.Y. Pretreatment with H2O2 alleviates aluminum-induced oxidative stress in wheat seedlings. J. Integr. Plant Biol. 2011, 53, 44–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Li, Y.; Wu, L.; Zhou, H.; Zhang, G.; Ma, Z. Ectopic expression of a novel Ser/Thr protein kinase from cotton (Gossypium barbadense), enhances resistance to Verticillium dahliae infection and oxidative stress in Arabidopsis. Plant Cell Rep. 2013, 32, 1703–1713. [Google Scholar] [CrossRef]
- Rossel, J.B.; Walter, P.B.; Hendrickson, L.; Chow, W.S.; Poole, A.; Mullineaux, P.M.; Pogson, B.J. A mutation affecting Ascorbate peroxidase 2 gene expression reveals a link between responses to high light and drought tolerance. Plant Cell Environ. 2006, 29, 269–281. [Google Scholar] [CrossRef]
- Prashanth, S.R.; Sadhasivam, V.; Parida, A. Over expression of cytosolic copper/zinc superoxide dismutase from a mangrove plant Avicennia marina in indica rice var Pusa Basmati-1 confers abiotic stress tolerance. Transgenic Res. 2008, 17, 281–291. [Google Scholar] [CrossRef]
- Badawi, G.H.; Kawano, N.; Yamauchi, Y.; Shimada, E.; Sasaki, R.; Kubo, A.; Tanaka, K. Over-expression of ascorbate peroxidase in tobacco chloroplasts enhances the tolerance to salt stress and water deficit. Physiol. Plant. 2004, 121, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Jiang, Y. Reactive oxygen species, antioxidant enzyme activities and gene expression patterns in leaves and roots of Kentucky bluegrass in response to drought stress and recovery. Sci. Hort. 2009, 120, 264–270. [Google Scholar] [CrossRef]
- Jiang, H.W.; Liu, M.J.; Chen, I.C.; Huang, C.H.; Chao, L.Y.; Hsieh, H.L. A glutathione S-transferase regulated by light and hormones participates in the modulation of Arabidopsis seedling development. Plant Physiol. 2010, 154, 1646–1658. [Google Scholar] [CrossRef] [PubMed]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Caninelli, A. Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Lamb, C.; Dixon, R. The oxidative burst in plant disease resistance. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef] [PubMed]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Savino, V.N.; Martelli, G.P.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the CoDiRO strain of Xylella fastidiosa subsp. pauca. BMC Genomics 2016, 1, 475. [Google Scholar] [CrossRef]
- Marra, F.P.; Marino, G.; Marchese, A.; Caruso, T. Effects of different irrigation regimes on a super-high-density olive grove cv. “Arbequina”: Vegetative growth, productivity and polyphenol content of the oil. Irrig. Sci. 2016, 34, 313–325. [Google Scholar] [CrossRef]
- Harper, S.J.; Ward, L.I.; Clover, G.R.G. Development of LAMP and Real-Time PCR Methods for the rapid detection of Xylella fastidiosa for quarantine and field applications. Phytopatology 2010, 100, 1282–1288. [Google Scholar] [CrossRef]
- Romanazzi, G.; Murolo, S.; Pizzichini, L.; Nardi, S. Esca in young and mature vineyards: And molecular diagnosis of the associated fungi. Eur. J. Plant Pathol. 2009, 125, 277–290. [Google Scholar] [CrossRef]
- Garrido, C.; Carbú, M.; Fernández-Acero, F.J.; Boonha, N.; Colyer, A.; Cantoral, J.M.; Budge, G. Development of protocols for detection of Colletotrichum acutatum and monitoring of strawberry anthracnose using qPCR. Plant Pathol. 2009, 58, 43–51. [Google Scholar] [CrossRef]
- Martín, M.T.; Cuesta, M.J.; Martín, L. Development of SCAR primers for PCR assay to detect Diplodia seriata. Int. Sch. Res. Not. 2014. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.T.; Cobos, R.; Martín, L.; López-Enríquez, L. qPCR Detection of Phaeomoniella chlamydospora and Phaeoacremonium aleophilum. Appl. Environ. Microb. 2012, 78, 3985–3991. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Raposo, R.; Lunello, P. A biomarker for the identification of four Phaeoacremonium species using the β-tubulin gene as the target sequence. Appl. Microbiol. Biot. 2008, 80, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Carlucci, A.; Lops, F.; Marchi, G.; Mugnai, L.; Surico, G. Has Xylella fastidiosa “Chosen” Olive Trees to Establish in the Mediterranean Basin? Phytopathol. Mediterr. 2013, 52, 541–544. [Google Scholar]
- Drenth, A.; Wagels, G.; Smith, B.; Sendall, B.; O’Dwyer, C.; Irvine, G.; Irwin, J.A.G. Development of a DNA-based method for detection and identification of Phytophthora species. Australas. Plant. Path. 2006, 35, 147–159. [Google Scholar] [CrossRef]
- Bilodeau, G.J.; Koike, S.T.; Uribe, P.; Martin, F.N. Development of an assay for rapid detection and quantification of Verticillium dahliae in soil. Phytopathology 2012, 102, 331–343. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimanting water deficits in leaves. Austral. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Rise, S.K. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol. 1977, 59, 304–309. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Spinach chloroplasts scavenge hydrogenperoxide on illumination. Plant Cell Physiol. 1980, 21, 1295–1307. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, S.K. Assay of catalase and peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Quackenbush, J. Microarray data normalization and transformation. Nat. Genet. 2002, 32, 496–501. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme activity (EU mg¯¹ protein) | ||||||
|---|---|---|---|---|---|---|
| APX | CAT | SOD | ||||
| Plant conditions | Cellina di Nardò | Leccino | Cellina di Nardò | Leccino | Cellina di Nardò | Leccino |
| Control | 3.20 ± 0.03 b, D | 3.66 ± 0.12 a, C | 4.82 ± 0.23 b, C | 5.23 ± 0.10 a, C | 12.94 ± 0.14 a, D | 13.46 ± 0.42 a, C |
| Drought | 4.34 ± 0.17 b, C | 9.23 ± 0.03 a, B | 8.53 ± 0.43 b, A | 10.73 ± 0.22 a, B | 17.64 ± 0.57 b, B | 27.02 ± 0.99 a, B |
| X.fastidiosa | 7.22 ± 0.07 b, A | 9.27 ± 0.12 a, B | 6.53 ± 0.32 b, B | 15.24 ± 0.39 a, A | 14.08 ± 0.20 b, C | 26.52 ± 0.17 a, B |
| Combined | 4.66 ± 0.10 b, B | 12.30 ± 0.13 a, A | 9.10 ± 0.19 b, A | 11.03 ± 0.03 a, B | 19.33 ± 0.24 b, A | 28.49 ± 0.25 a, A |
| Functional Categories and Genes | Abbr. | Primer | Sequence 5′-3′ | GeneBank |
|---|---|---|---|---|
| ROS scavenging activity genes related | ||||
| Ascorbate Peroxidase | APX | OeAPXF | CAAAAACTGCGCCCCTATAA | XM023040324.1 |
| OeAPXR | ACAGCAACAACACCAGCAAG | |||
| Catalase | CAT | OeCATF | GGATCCAGCCAGACAAGAGA | JQ429793 |
| OeCATR | TTGGCCTTACATTGAGACGA | |||
| Manganese Superoxide Dismutase | MnSOD | OeMnSODF | CTCCTGTTCGTGAAGGTGGT | AF427107 |
| OeMnSODR | GTGTCCAGACCAAGCCAAAT | |||
| Copper/Zinc Superoxide Dismutase | Cu/ZnSOD | OeCu/ZnSODF | CCATGCTGGTGATCTTGGTA | AF191342 |
| OeCu/ZnSODR | CAGTTCATGACCACCCCTTC | |||
| Pathogen genes related | ||||
| Leucine Rich Repeats | LRR-RLK | OeLRRF | CAACACAAGGCTTTTGGGACTT | XP_006367556.1 * |
| OeLRRR | TGTCATTGGTGCTTGTTGGT | |||
| Pathogenesis Protein Related | PR | OePRF | AACAAGGCTCGTGCAGAAGT | XM023013713 |
| OePRR | TCGACCCATGATCATAGCAA | |||
| Water deficit genes related | ||||
| Aquaporine | PIP2.1 | OePIP2.1F OePIP2.1R | TCTCGGGCCCTTGTTTTAGA AAAGAGAGGCCAGCAACCG | DQ202709 |
| Dehydration responsive element binding | DREB | OeDREBF OeDREBR | ACATGTTCTCCGCTCAGCTT GTGCCTCGTCTCCTTGAAAA | EF635424.1 |
| Dehydrin | DHN | OeDHNF | GGTTTGAAGGGGAAGGTTTC | KR349290.1 |
| OeDHNR | CTTCCTCAGCCTTCTTGTGG | |||
| Reference gene | ||||
| Polyubiquitin | UBQ | OeUBQF | GGTGGCCTCTAAATGTTCTTCTACTG | AF429430 * |
| OeUBQR | CACACAGACTTCATTAGAAAGACAATCA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Pascali, M.; Vergine, M.; Sabella, E.; Aprile, A.; Nutricati, E.; Nicolì, F.; Buja, I.; Negro, C.; Miceli, A.; Rampino, P.; et al. Molecular Effects of Xylella fastidiosa and Drought Combined Stress in Olive Trees. Plants 2019, 8, 437. https://doi.org/10.3390/plants8110437
De Pascali M, Vergine M, Sabella E, Aprile A, Nutricati E, Nicolì F, Buja I, Negro C, Miceli A, Rampino P, et al. Molecular Effects of Xylella fastidiosa and Drought Combined Stress in Olive Trees. Plants. 2019; 8(11):437. https://doi.org/10.3390/plants8110437
Chicago/Turabian StyleDe Pascali, Mariarosaria, Marzia Vergine, Erika Sabella, Alessio Aprile, Eliana Nutricati, Francesca Nicolì, Ilaria Buja, Carmine Negro, Antonio Miceli, Patrizia Rampino, and et al. 2019. "Molecular Effects of Xylella fastidiosa and Drought Combined Stress in Olive Trees" Plants 8, no. 11: 437. https://doi.org/10.3390/plants8110437