Distribution and Relative Abundance of Insect Vectors of Xylella fastidiosa in Olive Groves of the Iberian Peninsula

 ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
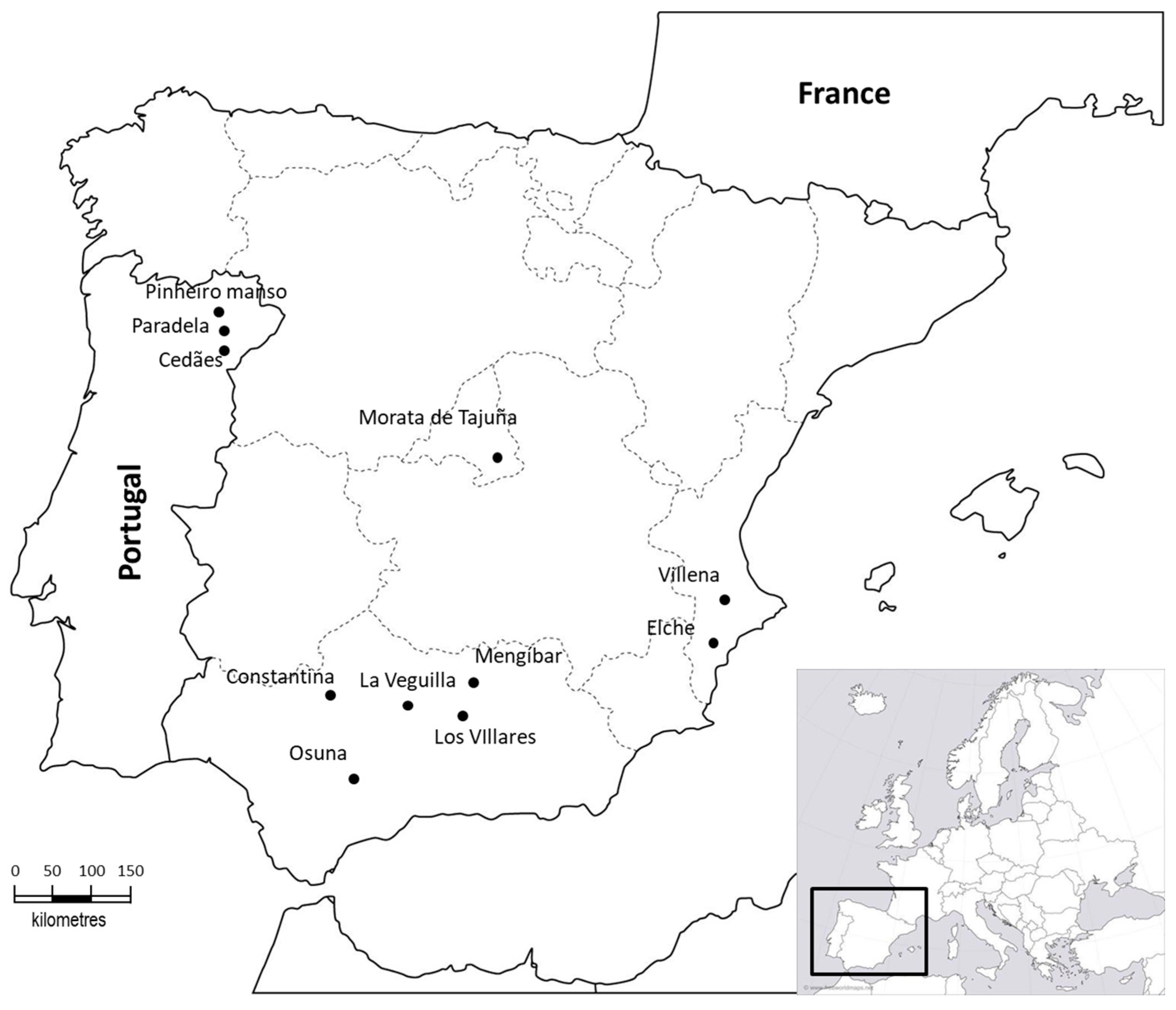
2.1. Field Sites
2.2. Sampling of Spittlebug Nymphs
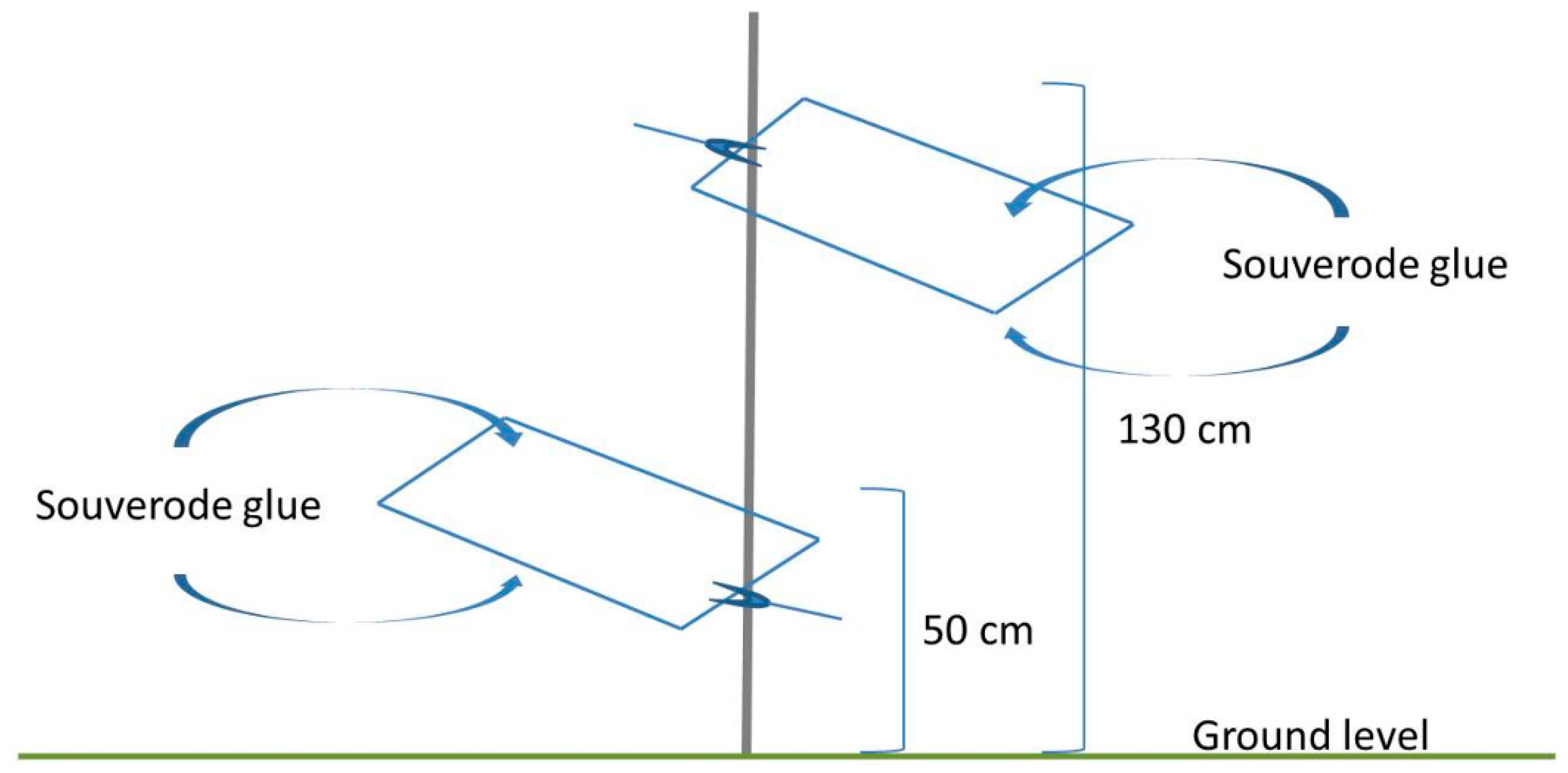
2.3. Sampling of Spittlebug Adults
2.3.1. Ground Vegetation
2.3.2. Olive Canopy
2.2.3. Unmanaged Oversummering Host Plants
3. Results
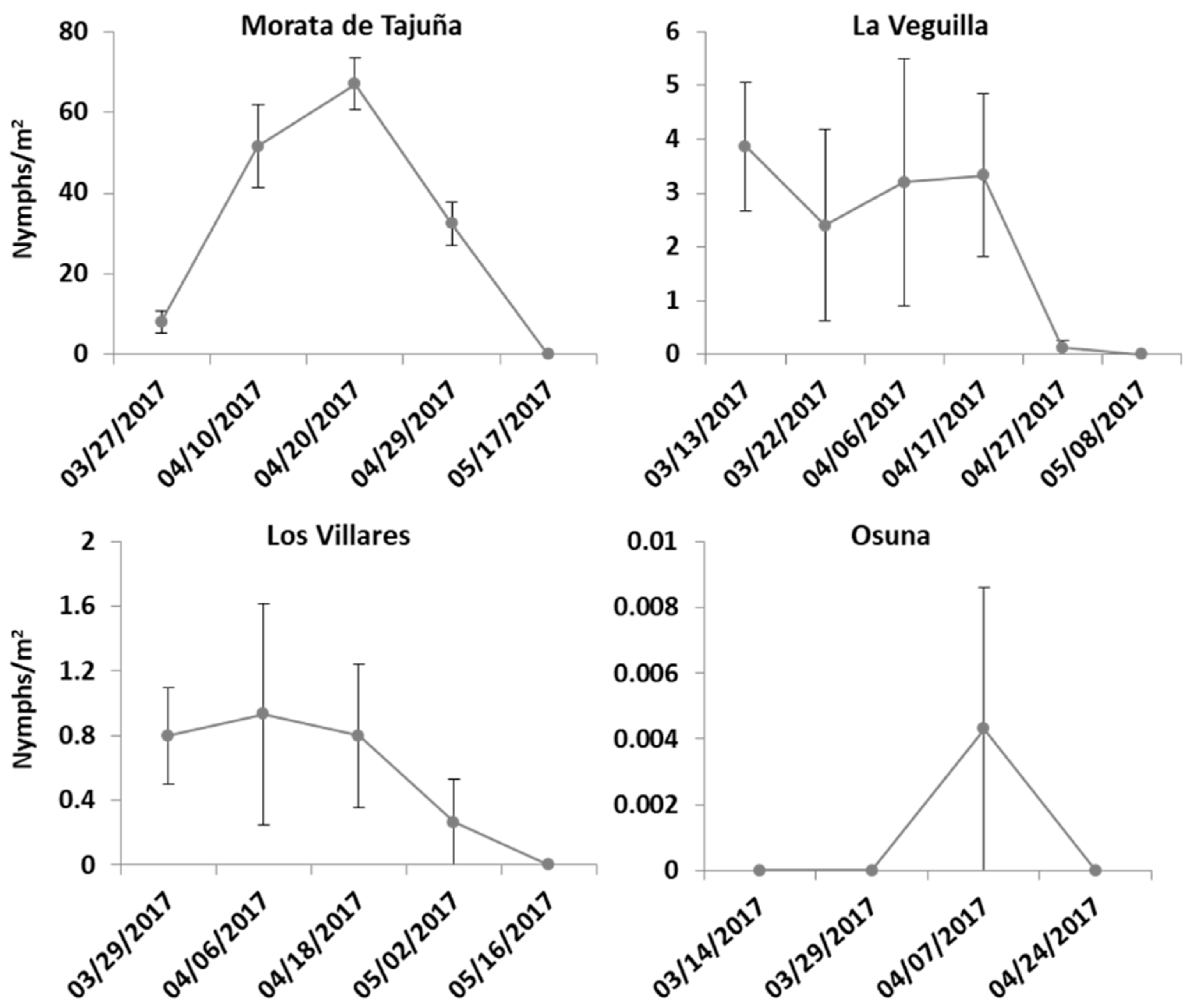
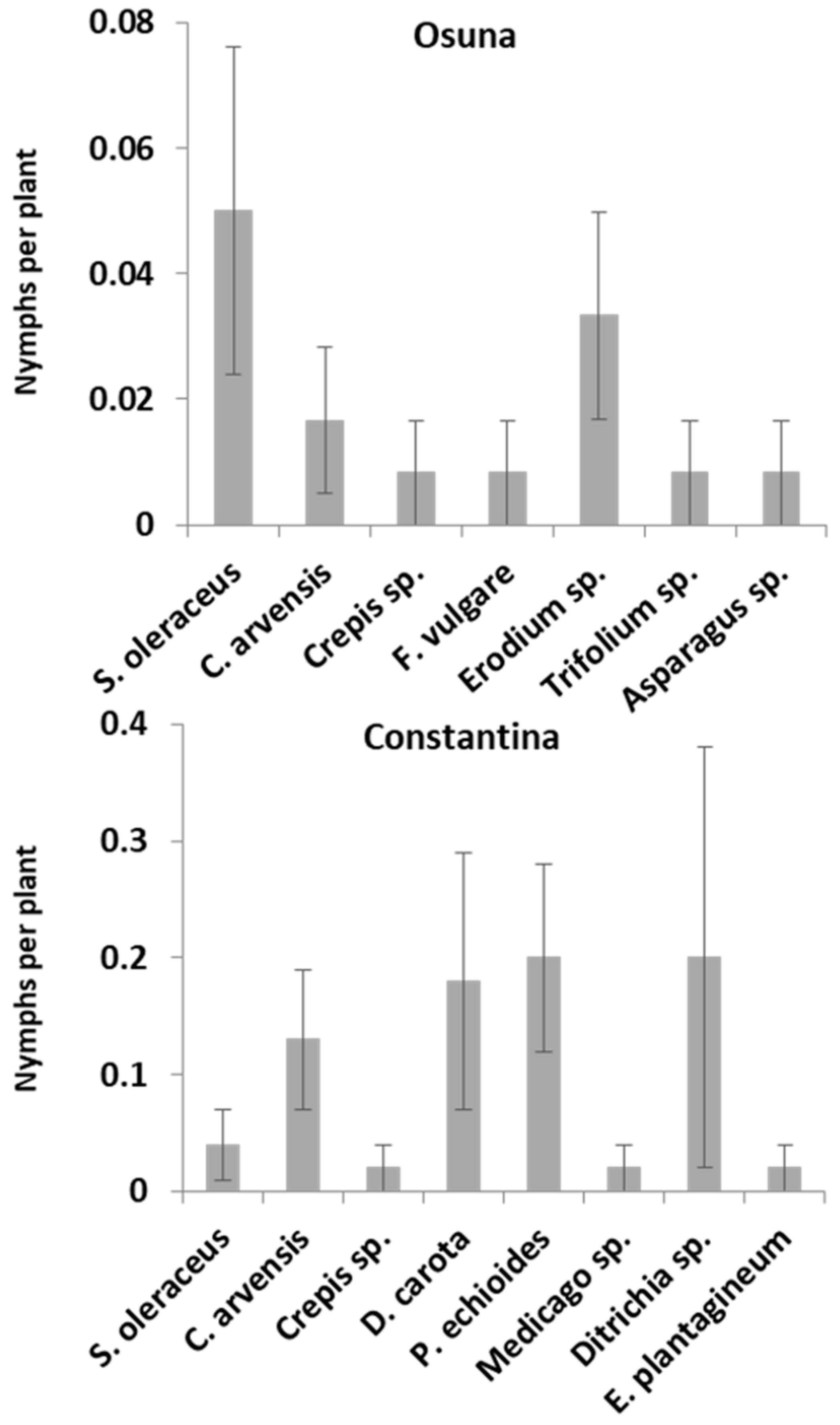
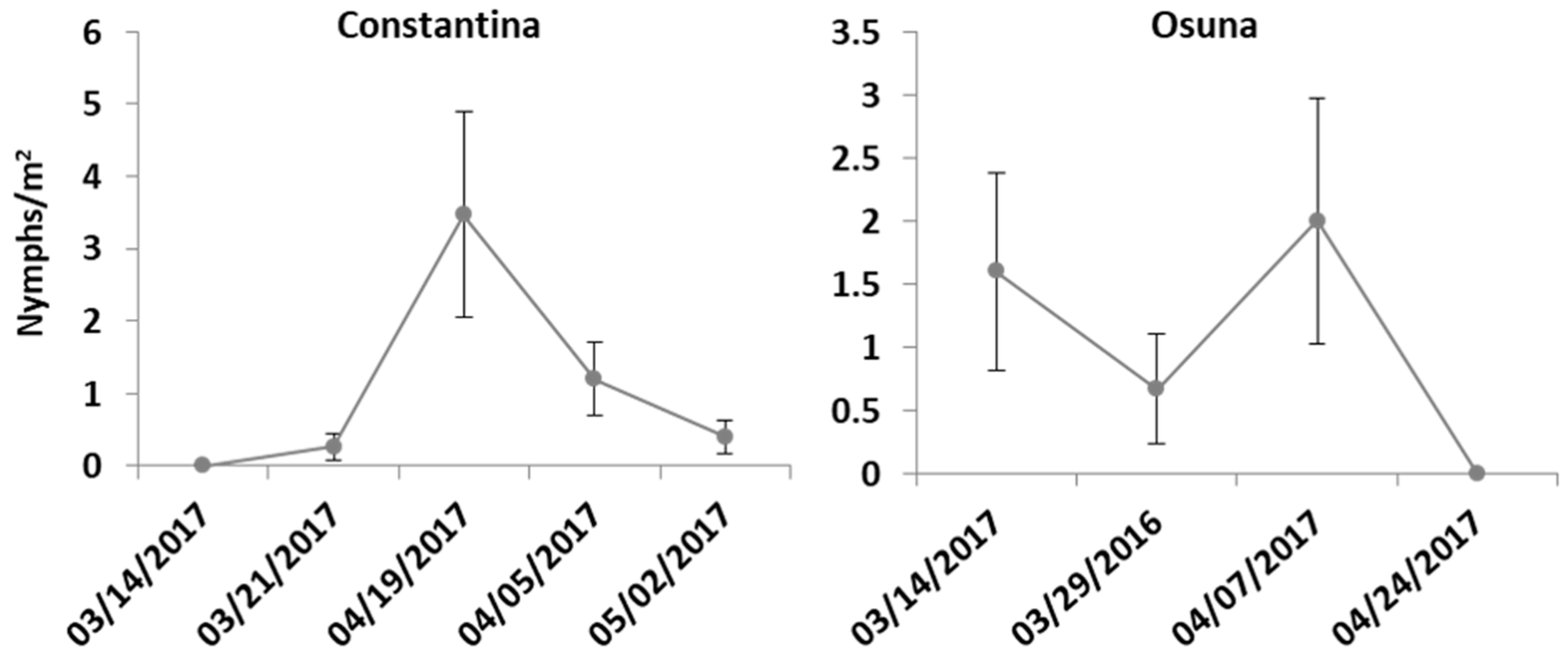
3.1. Population Abundance over Time and Host Plant Preference of Spittlebug Nymphs
3.2. Comparison of Different Sampling Methods to Monitor Spittlebug Populations
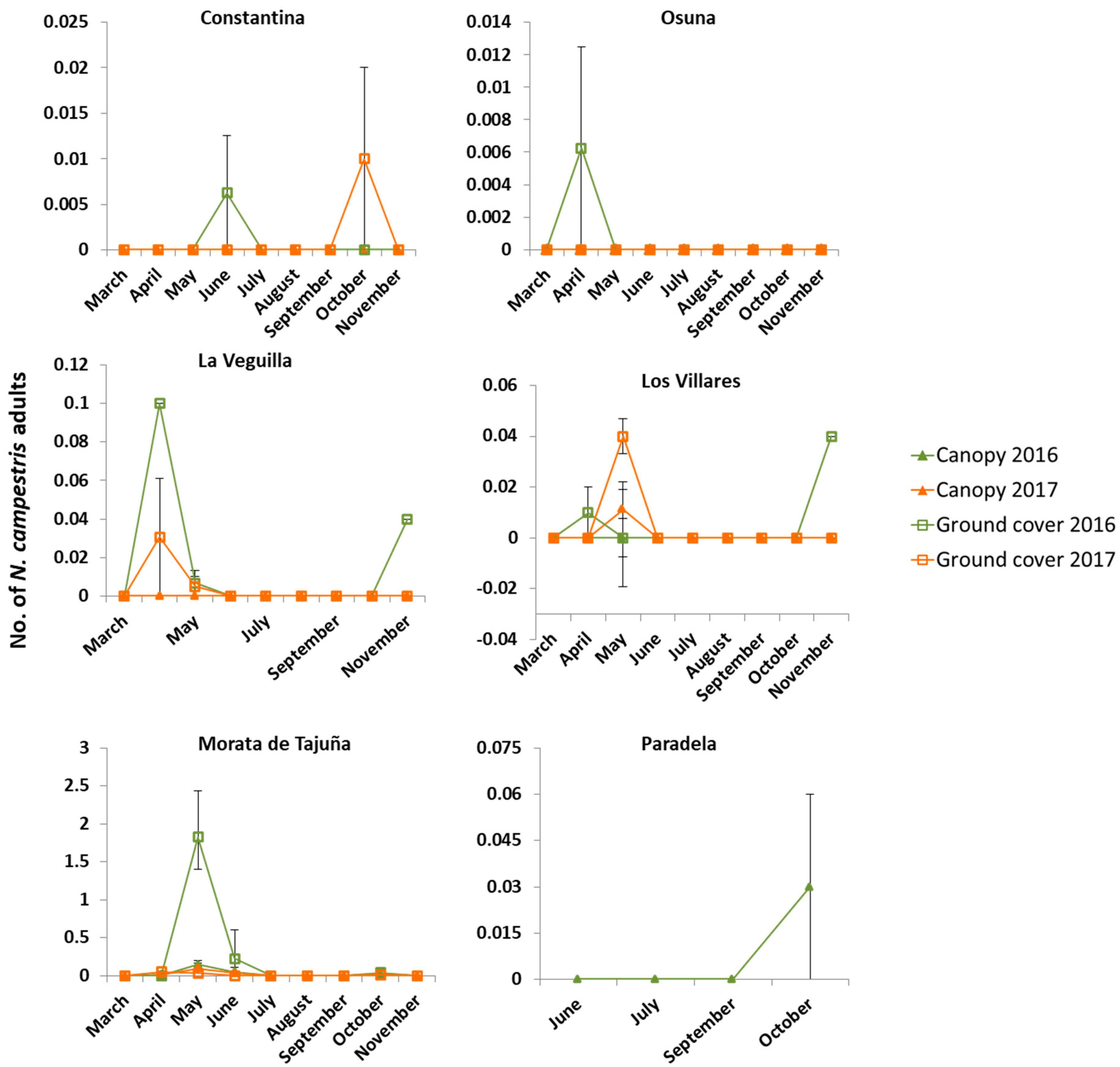
3.3. Distribution and Abundance over Time of Spittlebug Adults in the Ground Cover and Olive Canopy
3.4. Oversummering Hosts of Spittlebugs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Purcell, A.H. Xylella fastidiosa, a regional problem or global threat? J. Plant Pathol. 1997, 79, 99–105. [Google Scholar]
- Leu, L.S.; Su, C.C. Isolation, cultivation, and pathogenicity of Xylella fastidiosa, the causal bacterium of pear leaf scorch disease in Taiwan. Plant Dis. 1993, 77, 642–646. [Google Scholar] [CrossRef]
- Su, C.C.; Chang, C.J.; Chang, C.M.; Shih, H.T.; Tzeng, K.C.; Jan, F.J.; Deng, W.L. Pierce’s disease of grapevines in Taiwan: Isolation, cultivation and pathogenicity of Xylella fastidiosa. J. Phytopathol. 2013, 161, 389–396. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (southern Italy). J. Plant Pathol. 2013, 95, 659–668. [Google Scholar]
- Amanifar, N.; Taghavi, M.; Izadpanah, K.; Babaei, G. Isolation and pathogenicity of Xylella fastidiosa from grapevine and almond in Iran. Phytopatol. Mediterr. 2014, 53, 318. [Google Scholar] [CrossRef]
- Almeida, R.P.; Nunney, L. How Do Plant Diseases Caused by Xylella fastidiosa Emerge? Plant Dis. 2015, 99, 1457–1467. [Google Scholar] [CrossRef]
- Denancé, N.; Legendre, B.; Briand, M.; Olivier, V.; de Boisseson, C.; Poliakoff, F.; Jacques, M.-A. Several subspecies and sequence types are associated with the emergence of Xylella fastidiosa in natural settings in France. Plant Pathol. 2017, 66, 1054–1064. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Adrover, F.; Urbano, A.; Beidas, O.; Juan, A.; Montes-Borrego, M. First Detection of Xylella fastidiosa Infecting Cherry (Prunus avium) and Polygala myrtifolia Plants, in Mallorca Island, Spain. Plant Dis. 2017, 101, 1820. [Google Scholar] [CrossRef]
- EPPO (2018) EPPO Global Database. Available online: https://gd.eppo.int (accessed on 27 November 2018).
- Saponari, M.; Boscia, D.; Altamura, G.; Loconsole, G.; Zicca, S.; D’Attoma, G.; Morelli, M.; Palmisano, F.; Saponari, A.; Tavano, D.; et al. Isolation and pathogenicity of Xylella fastidiosa associated to the olive quick decline syndrome in southern Italy. Sci. Rep. 2017, 7, 17723. [Google Scholar] [CrossRef] [PubMed]
- MAPAMA. 2018. Available online: https://www.mapa.gob.es/es/ (accessed on 27 November 2018).
- Frazier, N.W. Xylem viruses and their insect vectors. In Proceedings of the International Conference on Virus and Vector on Perennial Hosts, Davis, CA, USA, 6–10 September 1965; pp. 91–99. [Google Scholar]
- Novotny, V.; Wilson, M.R. Why are there no small species among xylem-sucking insects? Evol. Ecol. 1977, 11, 419–437. [Google Scholar] [CrossRef]
- Purcell, A.H. Almond leaf scorch: Leafhopper and spittlebug vectors. J. Econ. Entomol. 1980, 73, 834–838. [Google Scholar] [CrossRef]
- Elbeaino, T.; Yaseen, T.; Valentini, F.; Ben Moussa, I.E.; Mazzoni, V.; D’onghia, A.M. Identification of three potential insect vectors of Xylella fastidiosa in southern Italy. Phytopatholol. Mediterr. 2014, 328–332. [Google Scholar] [CrossRef]
- Saponari, M.; Loconsole, G.; Cornara, D.; Yokomi, R.K.; De Stradis, A.; Boscia, D.; Porcelli, F. Infectivity and transmission of Xylella fastidiosa by Philaenus spumarius (Hemiptera: Aphrophoridae) in Apulia, Italy. J. Econ. Entomol. 2014, 107, 1316–1319. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.H.; Almeida, R.P. Insects as vectors of disease agents. Encycl. Plant Crop Sci. 2005, 10, 1–5. [Google Scholar]
- Rapicavoli, J.N. Structural and Functional Characterization of the Xylella fastidiosa O Antigen: Contribution to Bacterial Virulence, Insect Transmission, and Modulation of Innate Immune Recognition in Grapevine. Ph.D. Thesis, UC Riverside, Riverside, CA, USA, 2016. [Google Scholar]
- EFSA. Scientific opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2015, 13, 1–262. [Google Scholar] [CrossRef]
- Cornara, D.; Cavalieri, V.; Dongiovanni, C.; Altamura, G.; Palmisano, F.; Bosco, D.; Porcelli, F.; Almeida, R.P.P.; Saponari, M. Transmission of Xylella fastidiosa by naturally infected Philaenus spumarius (Hemiptera, Aphrophoridae) to different host plants. J. Appl. Entomol. 2016, 141, 80–87. [Google Scholar] [CrossRef]
- Cornara, D.; Saponari, M.; Zeilinger, A.R.; de Stradis, A.; Boscia, D.; Loconsole, G.; Porcelli, F. Spittlebugs as vectors of Xylella fastidiosa in olive orchards in Italy. J. Pest Sci. 2017, 90, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, V.; Dongiovanni, C.; Tauro, D.; Altamura, G.; Di Carolo, M.; Fumarola, G.; Saponari, M.; Bosco, D. Transmission of the codiro strain of Xylella fastidiosa by different insect species. In Proceedings of the XI European Congress of Entomology, Naples, Italy, 2–6 July 2018. [Google Scholar]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance become a new threat to European agriculture. J. Pest Sci. 2018, 1–16. [Google Scholar] [CrossRef]
- Cruaud, A.; Gonzalez, A.A.; Godefroid, M.; Nidelet, S.; Streito, J.C.; Thuillier, J.M.; Rasplus, J.Y. Using insects to detect, monitor and predict the distribution of Xylella fastidiosa: A case study in Corsica. Sci. Rep. 2018, 8, 15628. [Google Scholar] [CrossRef] [PubMed]
- Jacques, M.A.; Denancé, N.; Legendre, B.; Morel, E.; Briand, M.; Mississipi, S.; Crouzillat, D. New coffee plant-infecting Xylella fastidiosa variants derived via homologous recombination. Appl. Env. Microbiol. 2016, 82, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- Godefroid, M.; Cruaud, A.; Streito, J.C.; Rasplus, J.Y.; Rossi, J.P. Climate change and the potential distribution of Xylella fastidiosa in Europe. bioRxiv 2018. [Google Scholar] [CrossRef]
- Rathé, A.A.; Pilkington, L.J.; Gurr, G.M.; Hoddle, M.S.; Daugherty, M.P.; Constable, F.E.; Edwards, O.R. Incursion preparedness: Anticipating the arrival of an economically important plant pathogen Xylella fastidiosa Wells (Proteobacteria: Xanthomonadaceae) and the insect vector Homalodisca vitripennis (Germar) (Hemiptera: Cicadellidae) in Australia. Aust. Entomol. 2012, 51, 209–220. [Google Scholar] [CrossRef]
- Bonnier, G.; Layens, G.D.; Baulies, X. Claves Para la Determinación de Plantas Vasculares: Para Encontrar Fácilmente los Nombres de las Plantas sin Términos Técnicos; Omega: Barcelona, Spain, 1999. [Google Scholar]
- Dietrick, E.J. An improved backpack motor fan for suction sampling of insect populations. J. Econ. Entomol. 1961, 54, 394–395. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods; Blackwell Science Ltd.: Oxford, UK, 2000. [Google Scholar]
- Ruano, F.; Lozano, C.; Garcia, P.; Peña, A.; Tinaut, A.; Pascual, F.; Campos, M. Use of arthropods for the evaluation of the olive-orchard management regimes. Agric. For. Entomol. 2004, 6, 111–120. [Google Scholar] [CrossRef]
- Avinent, L.; Hermoso de Mendoza, A.; Llácer, G. Comparison of sampling methods to evaluate aphid populations (Homoptera, Aphidinea) alighting on apricot trees. Agronomie 1993, 13, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Ribaut, H. Faune de France. Homoptères Auchénorhynques. II. Iassidae 1952, 57, 1–474. [Google Scholar]
- Ossiannilsson, F. The Auchenorrhyncha (Homoptera) of Fennoscandia and Denmark. Part 2: The family Cicadidae, Cercopidae, Membracidae and Cicadellidae (excl. Deltocephalinae). Fauna Entomol. Scand. 1981, 7, 223–593. [Google Scholar]
- Della Giustina, W. Faune de France. France et Iles Anglo- Normandes. 73. Homopteres Cicadellidae. Volume 3. Complements aux Ouvrages a Henri Ribaut; Institut National de la Reserche Agronomique: Paris, France, 1989. [Google Scholar]
- Holzinger, W.; Kammerlander, I.; Nickel, H. The Auchenorrhyncha of Central Europe Die Zikaden Mitteleuropas Vol. 1 Fulgoromorpha, Cicadomorpha Excl; Cicadellidae; Brill Leiden: Boston, MA, USA, 2003; pp. 1–673. [Google Scholar]
- Mozaffarian, F.; Wilson, M.R. A checklist of the leafhoppers of Iran (Hemiptera: Auchenorrhyncha: Cicadellidae). Zootaxa 2016, 4062, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.R.; Landa, B.B.; Fereres, A. A survey of potential insect vectors of the plant pathogenic bacterium Xylella fastidiosa in three regions of Spain. Span. J. Agric. Res. 2014, 12, 795–800. [Google Scholar] [CrossRef]
- Purcell, A.H.; Gravena, S.; Donadio, L.C. Sharpshooter in citrus crops. Citrus-integrated management of insect and mite pests. Bebedouro Estação Exp. Citricul. 1994, 311, 213–229. [Google Scholar]
- Wilson, C.M.; Shade, R.E. Relative attractiveness of various luminescent colors to the cereal leaf beetle and the meadow spittlebug. J. Econ. Entomol. 1967, 60, 578–580. [Google Scholar] [CrossRef]
- Törmäla, T. Evaluation of five methods of sampling field layer arthropods, particularly the leafhopper community in grassland. Ann. Entomol. Fenicci. 1982, 48, 1–16. [Google Scholar]
- Tsagkarakis, A.E.; Afentoulis, D.G.; Matared, M.; Thanou, Z.N.; Stamatakou, G.D.; Kalaitzaki, A.P.; Perdikis, D.C. Identification and Seasonal Abundance of Auchenorrhyncha With a Focus on Potential Insect Vectors of Xylella fastidiosa in Olive Orchards in Three Regions of Greece. J. Econ. Entom. 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ben Moussa, I.E.; Mazzoni, V.; Valentini, F.; Yaseen, T.; Lorusso, D.; Speranza, S.; D’Onghia, A.M. Seasonal fluctuations of sap-feeding insect species infected by Xylella fastidiosa in Apulian olive groves of southern Italy. J. Econ. Entomol. 2016, 109, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.E.; Ruesink, W.G. Vector intensity: A product of propensity and activity. In Plant Virus Epidemics: Monitoring, Modelling and Predicting Outbreaks; McLean, G.D., Garrett, R.G., Ruesink, W.G., Eds.; Academic Press: Sydney, Australia, 1986. [Google Scholar]
- Weaver, C.R.; King, D.R. Meadow Spittlebug, Philaenus leucophthalmus (L.); Research Bulletin; Ohio Agricultural Experiment Station: Wooster, OH, USA, 1954. [Google Scholar]
- Nilakhe, S.S.; Buainain, C.M. Observations on movements of spittlebug adults. Pesquisa Agropecuaria Brasil. 1988, 23, 123–134. [Google Scholar]
- Dongiovanni, C.; Cavalieri, V.; Bodino, N.; Tauro, D.; Di Carolo, M.; Fumarola, G.; Altamura, G.; Lasorella, C.; Bosco, D. Plant selection and population trend of spittlebug immatures (Hemiptera: Aphrophoridae) in olive groves of the Apulia Region of Italy. J. Econ. Entomol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, V. Contribution to the knowledge of the Auchenorrhyncha (Hemiptera Fulgoromorpha and Cicadomorpha) of Tuscany (Italy). Redia 2005, 88, 85–102. [Google Scholar]
- Morris, M.G. Differences between the invertebrate faunas of grazed and ungrazed chalk grassland. IV. Abundance and diversity of Homoptera-Auchenorhyncha. J. Appl. Ecol. 1971, 37–52. [Google Scholar] [CrossRef]
- Drosopoulos, S.; Asche, M. Biosystematic studies on the spittlebug genus Philaenus with the description of a new species. Zool. J. Linn. Soc. 2001, 101, 169–177. [Google Scholar] [CrossRef]
- Almeida, R.P.P.; Purcell, A.H. Transmission of Xylella fastidiosa to Grapevines by Homalodisca coagulata (Hemiptera: Cicadellidae). J. Econ. Entomol. 2003, 96, 264–271. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spain | ||||||||
| Management | Constantina | Mengíbar | Los Villares | Morata de Tajuña | Osuna | La Veguilla | Villena (2016) | Elche (2017) |
| Herbicide | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 |
| Insecticide | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 |
| Mowing | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 |
| Grazing | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Portugal | ||||||||
| Management | Pinheiro Manso | Cedães | Paradela | |||||
| Herbicide | 0 | 0 | 1 | |||||
| Insecticide | 0 | 0 | 1 | |||||
| Mowing | 1 | 1 | 0 | |||||
| Grazing | 0 | 0 | 0 | |||||
| Plant Species | Presence of Philaenus spumarius | ||
|---|---|---|---|
| 21 June 2017 | 19 July 2017 | 7 September 2017 | |
| Anthemis arvensis (Compositae) | + | + | - |
| Carlina hispanica (Compositae) | + | + | - |
| Centaurea ornata (Compositae) | + | + | - |
| Cirsium arvense (Compositae) | + | + | - |
| Ditrichia sp. (Compositae) | - | - | - |
| Onopordum illyricum (Compositae) | + | + | - |
| Lavandula estoechas spp. pedunculata (Labiateae) | ++ | ++ | ++ |
| Thymus mastichina (Labiateae) | ++ | ++ | - |
| Daucus carota (Umbelliferae) | + | + | - |
| Echium plantagineum (Boraginaceae) | + | + | - |
| Avena sp. (Graminaeae) | ++ | ++ | - |
| Hypericum perforatum (Guttiferae) | + | + | - |
| Lomelosia sp. (Dipcasaceae) | + | + | - |
| Juniperus oxycedrus (Cupressaceae) | +++ | +++ | + |
| Quercus ilex (Fagaceae) | +++ | +++ | +++ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morente, M.; Cornara, D.; Plaza, M.; Durán, J.M.; Capiscol, C.; Trillo, R.; Ruiz, M.; Ruz, C.; Sanjuan, S.; Pereira, J.A.; et al. Distribution and Relative Abundance of Insect Vectors of Xylella fastidiosa in Olive Groves of the Iberian Peninsula. Insects 2018, 9, 175. https://doi.org/10.3390/insects9040175
Morente M, Cornara D, Plaza M, Durán JM, Capiscol C, Trillo R, Ruiz M, Ruz C, Sanjuan S, Pereira JA, et al. Distribution and Relative Abundance of Insect Vectors of Xylella fastidiosa in Olive Groves of the Iberian Peninsula. Insects. 2018; 9(4):175. https://doi.org/10.3390/insects9040175
Chicago/Turabian StyleMorente, Marina, Daniele Cornara, María Plaza, José Manuel Durán, Carmen Capiscol, Raquel Trillo, Manuel Ruiz, Carmen Ruz, Susana Sanjuan, Jose Alberto Pereira, and et al. 2018. "Distribution and Relative Abundance of Insect Vectors of Xylella fastidiosa in Olive Groves of the Iberian Peninsula" Insects 9, no. 4: 175. https://doi.org/10.3390/insects9040175